Различают постоянные и непостоянные (эпизодические) рефлекторные влияния на функциональное состояние дыхательного центра.
Пстоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Геринга - Брейера ), корня легкого и плевры (пульмоторакальный рефлекс ), хеморецепторов дуги аорты и сонных синусов (рефлекс Гейманса ), проприорецепторов дыхательных мышц.
Наиболее важным рефлексом является рефлекс Геринга - Брейера . В альвеолах легких заложены механорецепторы растяжения и спадения, являющиеся чувствительными нервными окончаниями блуждающего нерва. Любое увеличение объема легочных альвеол возбуждает эти рецепторы.
Рефлекс Геринга - Брейера является одним из механизмов саморегуляции дыхательного процесса, обеспечивая смену актов вдоха и выдоха. При растяжении альвеол во время вдоха нервные импульсы от рецепторов растяжения по блуждающему нерву идут к экспираторным нейронам, которые, возбуждаясь, тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает условия для повышения возбудимости инспираторной части дыхательного центра и осуществлению активного вдоха .
Кроме того, активность инспираторных нейронов усиливается при нарастании концентрации углекислого газа в крови, что также способствует проявлению вдоха.
Пульмоторакальный рефлекс возникает при возбуждении рецепторов, заложенных в легочной ткани и плевре. Проявляется этот рефлекс при растяжении легких и плевры. Рефлекторная дуга замыкается на уровне шейных и грудных сегментов спинного мозга.
К дыхательному центру постоянно поступают нервные импульсы от проприорецепторов дыхательных мышц. Во время вдоха происходит возбуждение проприорецепторов дыхательных мышц и нервные импульсы от них поступают в инспираторную часть дыхательного центра. Под влиянием нервных импульсов тормозится активность вдыхательных нейронов, что способствует наступлению выдоха.
Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждением разнообразных экстеро- и интерорецепторов . К ним относятся рефлексы, возникающие при раздражении рецепторов слизистой оболочки верхних дыхательных путей, слизистой носа, носоглотки, температурных и болевых рецепторов кожи, проприорецепторов скелетных мышц. Так, например, при внезапном вдыхании паров аммиака, хлора, сернистого ангидрида, табачного дыма и некоторых других веществ происходит раздражение рецепторов слизистой оболочки носа, глотки, гортани, что приводит к рефлекторному спазму голосовой щели, а иногда даже мускулатуры бронхов и рефлекторной задержке дыхания.
Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются ми-елинизированными волокнами блуждающего нерва.
Рефлекс Геринга - Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания.
У новорожденных рефлекс Геринга - Брейера четко проявляется только в первые 3-4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах.
Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через у-мотонейроны повышают активность о-мотонейронов и дозируют таким образом мышечное усилие.
Хеморефлексы дыхания. Рог и Рсог в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления Оз и выделение С02. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) - понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма 02, СОг и рН осуществляется периферическими и центральными хеморецепторами.
Адекватным раздражителем для периферических хеморецепторов является уменьшение Ро; артериальной крови, в меньшей степени увеличение Рсо2 и рН, а для центральных хеморецепторов - увеличение концентрации Н* во внеклеточной жидкости мозга.
Артериальные (периферические) хеморецепто-ры. Периферические хеморецепторы находятся в каротидных и
аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Ответ периферических хеморецепторов на понижение Рао^ является очень быстрым, но нелинейным. При Рао; в пределах 80-60мм рт. ст. (10,6-8,0 кПа) наблюдается слабое усиление вентиляции, а при Рао; ниже 50 мм рт. ст. (6,7 кПа) возникает выраженная гипервентиляция.
Расо2 и рН крови только потенцируют эффект гипоксии на артериальные хеморецепторы и не являются адекватными раздражителями для этого типа хеморецепторов дыхания.
Реакция артериальных хеморецепторов и дыхания на гипоксию. Недостаток С>2 в артериальной крови является основным раздражителем периферических хеморецепторов. Импульсная активность в афферентных волокнах синокаротидного нерва прекращается при Раод выше 400 мм рт. ст. (53,2 кПа). При нормоксии частота разрядов синокаротидного нерва составляет 10% от их максимальной реакции, которая наблюдается при Раод около 50 мм рт. ст. и ниже-Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их апаптации к высокогорью (3500 м и выше).
Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра.
Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции.
Адекватным раздражителемдля центральных хеморецепторов является изменение концентрации Н 4 во внеклеточной жидкости мозга.Функцию регулятора пороговыхсдвигов рН в области центральных хеморецепторов выполняют структуры гематоэнцефали-ческого барьера, который отделяет кровь отвнеклеточной жидкости мозга. Через этот барьер осуществляется транспорт02, С02 и Н^ между кровьюи внеклеточной жидкостью мозга.Транспорт СОз и H + из внутреннейсреды мозга в плазмукрови черезструктуры гематоэнцефалического барьерарегулируется с участием фермента ка рбоангидразы.
Реакция дыхания на COi. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга используют метод возвратного дыхания. Испытуемый дышит из замкнутой емкости, заполненной предварительно чистым Од. При дыхании в замкнутой
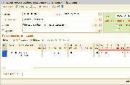
Рис. 8.12. Изменение вентиляциилегких (ve. л"мин) в зависимости от парциального давленияОд (А) иCOz <Б) в альвеолярном воздухе при различномсодержании Ог в альвеолярном воздухе (40, 50. 60 и 100 мм рт. ст.).
системе выдыхаемый СО; вызывает линейное увеличение концентрации СОа и одновременно повышает концентрацию Н* в крови, а также во внеклеточной жидкости мозга. Тест проводят в течение 4-5 мин под контролем содержания СО; в выдыхаемом воздухе-
На рис. 8.12 показано изменение объема вентиляции при различном уровне напряжения С02 в артериальной крови. При Расоа ниже 40 мм рт. ст. (5,3 кПа) может возникнуть апноэ в результате гипокапнии. В этот период дыхательный центр мало чувствителен к гипоксической стимуляции периферических хеморецепторов.
8.6.3. Координация дыхания с другими функциями организма
В филогенетическом развитии организма человека и животных дыхательный центр приобретает сложные синаптические взаимоотношения с различными отделами ЦНС.
В отличие отдругих физиологических функций организма дыханиенаходится под контролемавтономной (вегетативной) и соматическойнервной системы, поэтомуу человека и животных дыхание нередконазывают вегето-сомати ческой функцией.Существует тесное взаимодействиерегуляции дыхания гуморальной и рефлекторной природы и процессамисознательной деятельности мозга.Однако во время сна или в состояниях, связанных с отсутствиемсознания у человека,сохраняется внешнее дыхание и обеспечивается нормальное поддержаниегазового гомеостаза внутренней среды.С другой стороны, человек имеет возможность по собственному желанию
изменять глубину и частоту дыхания или задерживать его, например во время пребывания под водой. Произвольное управление дыханием основано на корковом представительстве проприоцептивного анализатора дыхательных мышц и на наличии коркового контроля дыхательных мышц.
Электрическое раздражение коры больших полушарий у человека и животных показало, что возбуждение одних корковых зон вызывает увеличение, а раздражение других - уменьшение легочной вентиляции. Наиболее сильное угнетение дыхания возникает при электрической стимуляции лимбической системы переднего мозга. При участии центров терморегуляции гипоталамуса возникает ги-перпноэ при гипертермических состояниях.
Однако многие нейрофизиологические механизмы взаимодействия нейронов переднего мозга с дыхательным центром остаются пока мало изученными.
Дыхание опосредованно черезгазы крови влияетна кровообращение вомногих органах.Важнейшим гуморальным, или метаболическим, регуляторомлокального мозгового кровотокаявляются Н* артериальнойкрови и межклеточной жидкости. Вкачестве метаболического регулятора тонусасосудов мозга рассматривают также С02. В последнеевремя этаточка зрения подвергается сомнению, поскольку СО-1 как молекулярноесоединение практически отсутствует во внутренней среде организма.Молекулярный С02 (0-С=-0) встречается в организмев альвеолярном воздухе, а в тканях только припереносе СОд через аэрогематический и гистогематический барьеры. В крови и межклеточной жидкости СО; находится в связанном состоянии, в видегидрокарбонатов, поэтому правильнее говорить о метаболической регуляции Н^ тонусагладких мышц артериальных сосудови их просвета. В головном мозгеповышение концентрации Н^ расширяет сосуды, апонижение концентрации I^ в артериальной крови или межклеточной жидкости, напротив, повышает тонусгладких мышц сосудистой стенки.Возникающие при этомизменения мозгового кровотока способствуютизменению градиента рН пообе стороны гематоэнцефалическогобарьера и создаютблагоприятные условиялибо для вымыванияиз сосудов мозга крови снизким значением рН,либо для понижения рН крови в результате замедления кровотока.
Функциональноевзаимодействие систем регуляции дыхания и кровообращенияявляется предметоминтенсивных физиологических исследований.Обе системы имеют общиерефлексогенные зоны в сосудах: аортальную и синокаротидные. Периферические хеморецепторы дыхания аортальныхи каротидных телец,чувствительные к гипоксии в артериальной крови, и барорецепторы стенки аорты и каротидных синусов,чувствительные к изменению системного артериального давления, расположены врефлексогенных зонах в непосредственной близостидруг от друга. Все названныерецепторы посылают афферентныесигналы к специализированнымнейронам основногочувствительного ядра продолговатого мозга -ядра одиночногопучка. В непосредственной близости от этогоядра находится
дорсальное дыхательное ядро дыхательного центра. Здесь же в продолговатом мозге находится сосудодвигательный центр.
Координацию деятельности дыхательного и сосудодвигательного центров продолговатого мозга осуществляют нейроны ряда интег-ративных ядер бульбарной ретикулярной формации.
Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны дыхательного центра имеют связи с многочисленными механорецепторами дыхательных путей и альвеол легких и рецепторов сосудистых рефлексогенных зон.
Рецепторы легких 1
В легких человека находятся следующие типы механорецепторов:
рецепторы растяжения гладких мышц дыхательных путей; Легочные рецепторы растяжения
ирритантные, или быстроадаптирующиеся, рецепторы слизистой оболочки дыхательных путей;
J-рецепторы.
Легочные рецепторы растяжения
Полагают, что эти рецепторы залегают в гладких мышцах воздухоносных путей.
Если легкие длительно удерживаются в раздутом состоянии, то активность рецепторов растяжения изменяется мало, что говорит об их слабой адаптируемости.
Импульсация от этих рецепторов идет по крупным миелиновым волокнам блуждающих нервов. Перерезка блуждающих нервов устраняет рефлексы с этих рецепторов.
Основной ответ на возбуждение легочных рецепторов растяжения - уменьшение частоты дыхания в результате увеличения времени выдоха. Эта реакция называется инфляционным рефлексом
Геринга - Брейера. (т. е. возникающим в ответ на раздувание)
В классических экспериментах показано, что раздувание легких приводит к затормаживанию дальнейшей активности инспираторных мышц.
Существует и обратная реакция, т. е. увеличение этой активности в ответ на уменьшеие объема легких (дефляционный рефлекс
). Эти рефлексы могут служить механизмом саморегуляции по принципу отрицательной обратной связи.
Когда-то считалось, что рефлексы Геринга-Брейера играют основную роль в регуляции вентиляции, т. е. именно от них зависит глубина и частота дыхания. Принцип такой регуляции мог бы заключаться в модулировании работы «прерывателя вдоха» в продолговатом мозге импульсацией от рецепторов растяжения. Действительно, при двустороннем пе^ ререзании блуждающих нервов у большинства животных устанавливается глубокое редкое дыхание. Однако в последних работах было показано, что у взрослого человека рефлексы Геринга-Брейера не действуют, пока дыхательный объем не превосходит 1 л (как, например, при физической нагрузке). Кратковременная двусторонняя блокада блуждающих нервов посредством местной анестезии у бодрствующего человека не влияет ни на частоту, ни на глубину дыхания. Некоторые данные указывают, что эти рефлексы могут иметь большее значение для новорожденных.
Рефлексы со слизистой оболочки полости носа.
Раздражение ирритантных рецепторов слизистой оболочки полости носа, например табачным дымом, инертными частицами пыли, газообразными веществами, водой вызывает сужение бронхов, голосовой щели, брадикардию, снижение сердечного выброса, сужение просвета сосудов кожи и мышц. Защитный рефлекс проявляется у новорожденных при кратковременном погружении в воду. У них возникает остановка дыхания, препятствующая проникновению воды в верхние дыхательные пути.
Рефлексы с глотки. Механическое раздражение рецепторов слизистой оболочки задней части полости носа вызывает сильнейшее сокращение диафрагмы, наружных межреберных мышц, а следовательно, вдох, который открывает дыхательный путь через носовые ходы (аспирационный рефлекс). Этот рефлекс выражен у новорожденных.
Рефлексы с гортани и трахеи.
Многочисленные нервные окончания расположены между эпителиальными клетками слизистой оболочки гортани и главных бронхов. Эти рецепторы раздражаются вдыхаемыми частицами, раздражающими газами, бронхиальным секретом, инородными телами. Все это вызывает кашлевой рефлекс, проявляющийся в резком выдохе на фоне сужения гортани и сокращение гладких мышц бронхов, которое сохраняется долгое время после рефлекса.
Кашлевой рефлекс является основным легочным рефлексом блуждающего нерва.
Рефлексы с рецепторов бронхиол.
Многочисленные миелинизированные рецепторы находятся в эпителии внутрилегочных бронхов и бронхиол. Раздражение этих рецепторов вызывает гиперпноэ, бронхоконстрикцию, сокращение гортани, гиперсекрецию слизи, но никогда не сопровождается кашлем.
Рецепторы наиболее чувствительны к трем типам раздражителей: 1) табачному дыму, многочисленным инертным и раздражающим химическим веществам;
2) повреждению и механическому растяжению дыхательных путей при глубоком дыхании, а также пневмотораксе, ателектазах, действии бронхоконстрикторов;
3) легочной эмболии, легочной капиллярной гипертензии и к легочным анафилактическим феноменам.
Рефлексы с J-рецепторов
. В альвеолярных перегородках в контакте с капиллярами находятся особые J-рецепторы. Эти рецепторы особенно чувствительны к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам, фенилдигуаниду (при внутривенном введении этого вещества). Стимуляция J-рецепторов вызывает вначале апноэ, затем поверхностное тахипноэ, гипотензию и брадикардию.
Рефлексы Геринга - Брейера.
Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются ми-елинизированными волокнами блуждающего нерва.
Рефлекс Геринга - Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания.
У новорожденных рефлекс Геринга - Брейера четко проявляется только в первые 3-4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах.
Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через у-мотонейроны повышают активность о-мотонейронов и дозируют таким образом мышечное усилие.
Хеморефлексы дыхания. Рог и Рсог в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления Оз и выделение С02. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) - понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма 02, СОг и рН осуществляется периферическими и центральными хеморецепторами.
Адекватным раздражителем для периферических хеморецепторов является уменьшение Ро; артериальной крови, в меньшей степени увеличение Рсо2 и рН, а для центральных хеморецепторов - увеличение концентрации Н* во внеклеточной жидкости мозга.
Артериальные (периферические) хеморецепто-ры. Периферические хеморецепторы находятся в каротидных и
аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Ответ периферических хеморецепторов на понижение Рао^ является очень быстрым, но нелинейным. При Рао; в пределах 80-60мм
рт. ст. (10,6-8,0 кПа) наблюдается слабое усиление вентиляции, а при Рао; ниже 50 мм рт. ст. (6,7 кПа) возникает выраженная гипервентиляция.
Расо2 и рН крови только потенцируют эффект гипоксии на артериальные хеморецепторы и не являются адекватными раздражителями для этого типа хеморецепторов дыхания.
Реакция артериальных хеморецепторов и дыхания на гипоксию. Недостаток С>2 в артериальной крови является основным раздражителем периферических хеморецепторов. Импульсная активность в афферентных волокнах синокаротидного нерва прекращается при Раод выше 400 мм рт. ст. (53,2 кПа). При нормоксии частота разрядов синокаротидного нерва составляет 10% от их максимальной реакции, которая наблюдается при Раод около 50 мм рт. ст. и ниже-Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их апаптации к высокогорью (3500 м и выше).
Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра.
Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции.
Адекватным раздражителемдля центральных хеморецепторов является изменение
концентрации Н 4 во внеклеточной жидкости мозга.Функцию
регулятора пороговыхсдвигов рН в области
центральных хеморецепторов выполняют структуры гематоэнцефали-ческого барьера, который отделяет кровь отвнеклеточной жидкости мозга.
Через этот барьер осуществляется транспорт02, С02 и Н^
между кровьюи внеклеточной
жидкостью мозга.Транспорт СОз и H + из
внутреннейсреды мозга в
плазмукрови
черезструктуры гематоэнцефалического
барьерарегулируется с участием фермента ка рбоангидразы.
50. Регуляция дыхания при пониженном и повышенном атмосферном давлении.
Дыхание при пониженном атмосферном давлении. Гипоксия
Атмосферное давление понижается при подъеме на высоту. Это сопровождается одновременным снижением парциального давления кислорода в альвеолярном воздухе. На уровне моря оно составляет 105 мм.рт.ст. На высоте 4000 м уже в 2 раза меньше. В результате уменьшается напряжение кислорода в крови. Возникает гипоксия. При быстром падении атмосферного давления наблюдается острая гипоксия. Она сопровождается эйфорией, чувством ложного благополучия, и скоротечной потерей сознания. При медленном подъеме гипоксия нарастает медленно. Развиваются симптомы горной болезни. Первоначально появляется слабость, учащение и углубление дыхания, головная боль. Затем начинаются тошнота, рвота, резко усиливаются слабость и одышка. В итоге также наступает потеря сознания, отек мозга и смерть. До высоты 3 км у большинства людей симптомов горной болезни не бывает. На высоте 5 км наблюдаются изменения дыхания, кровообращения, высшей нервной деятельности. На высоте 7 км эти явления резко усиливаются. Высота 8 км является предельной для жизнедеятельности высоте организм страдает не только от гипоксии, но и от гипокапнии. В результате снижения напряжения кислорода в крови возбуждаются хеморецепторы сосудов. Дыхание учащается и углубляется. Из крови выводится углекислый газ и его напряжение падает ниже нормы. Это приводит к угнетению дыхательного центра. Несмотря на гипоксию дыхание становится редким и поверхностным. В процессе адаптации к хронической гипоксии выделяют три стадии. На первой, аварийной, компенсация достигается за счет увеличения легочной вентиляции, усиления кровообращения, повышения кислородной емкости крови и т.д. На стадии относительной стабилизации происходят такие изменения систем, организма, которые обеспечивают более высокий, и выгодный уровень адаптации. В стабильной стадии физиологические показатели организма становятся устойчивыми за счет ряда компенсаторных механизмов. Так кислородная емкость крови увеличивается не только за счет возрастания количества эритроцитов, но и 2,3-фосфоглицерата в них. За счет 2,3-фосфоглицерата улучшается диссоциация оксигемоглобина в тканях. Появляется фетальный гемоглобин, имеющий более высокую способность связывать кислород. Одновременно повышается диффузионная способность легких и возникает "функциональная эмфизема". Т.е. в дыхание включаются резервные альвеолы и увеличивается функциональная остаточная емкость. Энергетический обмен понижается, но повышается интенсивность обмена углеводов.
Гипоксия это недостаточное снабжение тканей кислородом. Формы гипоксии:
1. Гипоксемическая гипоксия. Возникает при снижении напряжения кислорода в крови (уменьшение атмосферного давления, диффузионной способности легких и т.д.).
2. Анемическая гипоксия. Является следствием понижения способности крови транспортировать кислород (анемии, угарное отравление).
3. Циркуляторная гипоксия. Наблюдается при нарушениях системного и местного кровотока (болезни сердца и сосудов).
4. Гистотоксическая гипоксия. Возникает при нарушении тканевого дыхания (отравление цианидами).
Дыхание человека при повышенном давлении воздуха имеет место на значительной глубине под водой при работе водолазов или при кессонных работах. Поскольку давление одной атмосферы соответствует давлению столба воды высотой 10 м, то в соответствии с глубиной погружения человека под воду в скафандре водолаза или в кессоне поддерживается давление воздуха по этому расчету. Человек, находясь в атмосфере повышенного давления воздуха, не испытывает каких-либо дыхательных расстройств. При повышенном давлении атмосферного воздуха человек может дышать в том случае, если в его дыхательные пути поступает воздух под таким же давлением. При этом растворимость газов в жидкости прямо пропорциональна его парциальному давлению.
Поэтому при дыхании воздухом на уровне моря в 1 мл крови содержится 0,011 мл физически растворенного азота. При давлении воздуха, которым дышит человек, например, 5 атмосфер, в 1 мл крови будет содержаться в 5 раз больше физически растворенного азота. При переходе человека к дыханию при более низком давлении воздуха (подъем кессона на поверхность или всплытие водолаза) кровь и ткани тела могут удержать только 0,011 мл N2/мл крови. Остальное количество азота переходит из раствора в газообразное состояние. Переход человека из зоны повышенного давления вдыхаемого воздуха к более низкому его давлению должен происходить достаточно медленно, чтобы освобождающийся азот успел выделиться через легкие. Если азот, переходя в газообразное состояние, не успевает полностью выделиться через легкие, что имеет место при быстром подъеме кессона или нарушении режима всплытия водолаза, пузырьки азота в крови могут закупорить мелкие сосуды тканей организма. Это состояние называется газовая эмболия. В зависимости от локализации газовой эмболии (сосуды кожи, мышц, центральной нервной системы, сердца и др.) у человека возникают различные расстройства (боли в суставах и мышцах, потеря сознания), которые в целом называются «кессонной болезнью».
Брейер Йозеф - австралийский врач и физиолог, которого Зигмунд Фрейд и другие называли родоначальником психоанализа. Ему удалось излечить больную от симптомов истерии после того как он под гипнозом помог ей вспомнить неприятные моменты из прошлого. Он рассказал о своем методе и полученных результатах Зигмунду Фрейду, а также передал ему своих пациентов.
Йозеф Брейер: биография
Родился 15.01.1842 г. в Вене и умер там же 20.06.1925. Отец Йозефа Леопольд (1791-1872) был преподавателем религии, нанятым венской еврейской общиной. Брейер описывал его как принадлежащего к «тому поколению восточноевропейских евреев, которое первым вышло из интеллектуального гетто на воздух западного мира».
Мать умерла, когда ему было около четырех лет, и ранние годы жизни Брейер Йозеф провел с бабушкой. Отец обучал его до восьми, а затем он поступил в Академическую гимназию Вены, которую окончил в 1858 г. В следующем году, пройдя курс общеуниверситетского образования, Йозеф Брейер поступил в медицинскую школу Венского университета и окончил медицинское обучение в 1867 году. В том же году, сразу после сдачи экзамена, он стал помощником терапевта Иоганна Оппольцера. Когда тот умер в 1871 году, Брейер начал собственную частную практику.
Лучший врач Вены
В 1875 году Брейер стал приват-доцентом терапии. Он ушел с этой должности 7 июля 1885 г., так как ему было отказано в доступе к пациентам с учебными целями. Также он отказался позволить хирургу Бильроту выдвинуть его на звание адъюнкт-профессора. Его формальные отношения с медицинским факультетом были, таким образом, напряженными.
В то же время Брейер был признан одним из лучших врачей и ученых Вены. Работа стала его главным интересом, и хотя он однажды назвал себя «врачом общей практики», он являлся, что сегодня называется, терапевтом. Некоторое представление о репутации Брейера может дать тот факт, что среди его пациентов были многие профессора медицинского факультета, а также Зигмунд Фрейд и премьер-министр Венгрии. В 1894 году он был избран в венскую Академию наук по представлению ее самых выдающихся членов: физика Эрнста Маха и физиологов Эвальда Геринга и Зигмунда Экснера.

Личная жизнь
20 мая 1868 г. Брейер Йозеф женился на Матильде Альтман, которая родила ему пятерых детей: Хаммершляг, Маргарет Шифф, Ганса и Дору. Дочь Брейера Дора совершила самоубийство, не желая быть схваченной нацистами. Они же убили внучку Брейера Ханну Шифф. Остальные его потомки живут в Англии, Канаде и Соединенных Штатах.
Научная работа
Брейер Йозеф изучал медицину в Вене и получил ученую степень в 1864 г. Он исследовал терморегуляцию и физиологию дыхания (рефлекс Геринг-Брейера). В 1871 году начал свою практику в Вене. Одновременно проводил исследования функции внутреннего уха (Теория Маха-Брейера о течении эндолимфальной жидкости). Став терапевтом в 1874 году, он вернулся к исследованиям в 1884-м.
Брейер был другом и семейным врачом некоторых членов Венского педагогического колледжа и столичного высшего общества. Он поддерживал переписку с художниками, писателями, философами, психологами и коллегами в своей области, а в 1894 году был избран членом-корреспондентом Академии наук.
Хорошо разбиравшийся в философии, Брейер Йозеф интересовался теорией познания и теоретическими основами дарвинизма, что подтверждается его участием в конференции 1902 г. и обменом письмами с Францем фон Брентано. Он был активным участником дискуссий об основах политики и идеологии, а также обсуждал вопросы искусства, литературы и музыки.
Как ассимилированный и просвещенный еврей, он принял своего рода пантеизм, перенятый им у Гете и Густава Теодора Фехнера. Его любимым афоризмом было высказывание Спинозы Suum esse conservare («Сохранять свое существование»). Он был охвачен формой скептицизма и, следуя Уильяму Теккерею, «демоном «но», что вынудило его подвергать сомнению любые вновь приобретенные знания. Из-за детального знания истории идей, социальной истории и политических условий своей эпохи, а также по причинам, связанным с его собственной жизнью, он считал, что для него было почти невозможным предпринять сомнительные действия.
В основе исследования Брейера в области физиологии было стремление найти взаимосвязь между структурой и функцией, и, следовательно, выявить форму телеологического запроса. Он интересовался регуляторными процессами в виде механизмов самоконтроля. В отличие от ряда физиологов в так называемом движении биофизикалистов, вдохновленный Эрнстом Брюкке, Германом фон Гельмгольцем и Дюбуа-Реймоном, Брейер верил в неовитализм.

Начала психоанализа
В 1880-1882 годах он лечил молодую пациентку Берту Паппенгейм (Анну О.), которая страдала нервным кашлем и множеством других истерических симптомов (перепадами настроения, изменениями в состояниях сознания, нарушениями зрения, параличом и судорогами, афазией). Во время долгих бесед врач и его подопечная увидели, что некоторые проявления болезни исчезли, когда были восстановлены воспоминания об их первом проявлении, и стало возможным воспроизвести связанные с ними аффекты. Это происходило в определенное время суток при спонтанных автогипнотических состояниях. На основании этих наблюдений, изначально случайно, пациент и врач разработали систематическую процедуру, согласно которой отдельные симптомы постепенно вспоминались в обратном хронологическом порядке, пока они не исчезли после полного воспроизведения исходной сцены. Иногда во время терапии использовался искусственный гипноз, если пациент не входил в состояние самогипноза.
Во время проведения терапии требовалось постоянное пребывание Анны О. в клинике недалеко от Вены из-за повышенного риска самоубийства больной. Несмотря на явный и неожиданный успех метода, некоторые проявления болезни оставались. Они включали временное забывание родного языка и сильнейшую невралгию тройничного нерва, которая требовала лечения морфином, вызывающим привыкание. Из-за этих симптомов Брейер направил пациентку на дальнейшее лечение к доктору Людвигу Бинсвангеру в санаторий Bellevue в Кройцлингене в июле 1882 года. Она выписалась в октябре с улучшениями, но полностью не излечившись.

Совместная работа с Фрейдом
В 1882 году Брейер Йозеф обсуждал вышеописанный случай со своим коллегой Зигмундом Фрейдом, который был на 14 лет его моложе. После того как последний начал работать невропатологом, он испытал данный метод на своих пациентах. Исходя из теории Шарко, Пьера Жане, Мёбиуса, Ипполита Бернхайма и других, они совместно разработали теоретические основы функционирования психического аппарата, а также терапевтические процедуры, которые они назвали «методом катарсиса», ссылаясь на идеи Аристотеля о функции трагедии (катарсис как очищение эмоций зрителей).
В 1893 году они опубликовали предварительный доклад «О психических механизмах истерических явлений». Два года спустя за ним последовали «Исследования истерии», «краеугольный камень психоанализа», заложивший основы в данной области психиатрии. В работе была глава, посвященная теории (Брейер), еще одна, посвященная терапии (Фрейд), и пять историй болезни (Анна О., Эмми фон Н., Катарина, Люси Р., Элизабет фон Р.).

Уход из психоанализа
Фрейд продолжал развивать теорию и технику во время совместной работы с Брейером (защитные неврозы, свободные ассоциации). Йозеф не был убежден в необходимости исключительной акцентации сексуальных факторов, и его коллега увидел в этом предостережении признак отстраненности. В 1895 году дистанция между ними увеличилась, что привело к концу их сотрудничества.
Продолжая проявлять интерес к развитию Брейер Йозеф катартический метод отбросил. Фрейд позже предложил гипотезу о том, что лечение Анны О. было внезапно прервано из-за сильного эротического переноса, сопровождаемого истерической беременностью и родами. Эта версия событий, воссозданная Фрейдом и распространенная Эрнестом Джонсом, среди прочего, не выдерживает исторической критики. Более поздние попытки показать, что описание случая Анны О. было мошенничеством, фактами подкреплены не были.

Разносторонняя личность
Йозеф Брейер был дружен со многими ярчайшими интеллектуалами своего времени. Он вел долгую переписку с Брентано, был близким другом поэтессы Марии фон Эбнер-Эшенбах и дружил с Махом, с которым познакомился во время исследований внутреннего уха. Мнение Брейера по литературным и философским вопросам, по-видимому, пользовалось широким уважением. Брейер владел многими языками: например, лечение Анны О. длительное время велось на английском языке. Диапазон и глубина его культурных интересов были столь же необычными и важными, как и его медицинские и научные достижения.

Йозеф Брейер: интересные факты из жизни
- После того как у его пациентки Анны О. сформировалась к нему сильная привязанность, носившая выраженный сексуальный характер, Брейер Йозеф работы в области психотерапии, требовавшие непосредственного контакта с пациентами, переложил на Зигмунда Фрейда.
- Брейер открыл, что невротические симптомы возникают по причине подсознательных процессов, и уходят, когда они становятся осознанными.
- Зигмунд Фрейд достижениями в психотерапии обязан Брейеру, который познакомил его со своими открытиями и передал ему своих пациентов.
- В 1868 г. он описал рефлекс Геринга-Брейера, который задействован для контроля вдыханий и вдыханий в процессе нормального дыхания.
- В 1873 г. Брейер открыл сенсорную функцию полукружных каналов костного лабиринта внутреннего уха и их связь с ориентацией в пространстве и с чувством равновесия.
- В своем завещании он выразил желание быть кремированным, и оно было исполнено.
Значение блуждающих нервов в саморегуляции дыхательного цикла установили Геринг и Брейер в опыте с раздуванием легких воздухом в различные фазы дыхательного цикла - раздувание легких воздухом тормозит вдох, после чего наступает выдох. Уменьшение объема легких (забор воздуха) тормозит выдох, ускоряет вдох. После перерезки блуждающих нервов раздувание легких не изменяет характер дыхания. Во время вдоха вследствие растяжения легких возбуждаются их механорецепторы (рецепторы растяжения - они локализуются в стенке трахеи и бронхов). Афферентные импульсы по блуждающим нервам поступают к дыхательным нейронам, тормозят вдох и способствуют смене вдоха на выдох (рефлекс Геринга-Брейера). При этом возбуждаются экспираторные и поздние инспираторные нейроны, которые, в свою очередь, тормозят ранние инспираторные нейроны.
Импульсация от проприорецепторов усиливает сокращение дыхательной мускулатуры и способствует смене вдоха на выдох. Причем главную роль играют мышечные и сухожильные рецепторы межреберных мышц и мышц брюшной стенки, которые содержат большое количество этих рецепторов.
Влияние интеро- и экстерорецептив- ных рефлексогенных зон на дыхание
Возбуждение рецепторов верхних дыхательных путей (они в основном холодовые) оказывает слабое тормозное влияние на дыхание.
Раздражение обонятельных рецепторов пахучими веществами в умеренной концентрации вызывает короткие вдохи - принюхивание. Однако адекватных рецепторов, воспринимающих изменение содержания 0 2 и С0 2 в воздухе и обеспечивающих возникновение соответствующих ощущений, у человека нет. И все же люди чувствуют уменьшение содержания кислорода в газовых смесях: некоторые отмечают снижение 0 2 в газовой смеси уже до уровня 12 % и почти все - до 9 %. Человек испытывает также затруднения при дыхании газовыми смесями с повышенным содержанием С0 2 .
Сильное раздражение слизистых оболочек воздухоносных путей (пыль, едкие пары, например, аммиака и инородные тела) вызывает возбуждение окончаний тройничного нерва - при этом возникает чиханье, возможно апноэ (остановка дыхания).
J-рецепторы (рецепторы интер- стиция) возбуждаются при скоплении жидкости в альвеолярной стенке (отек) и под влиянием ряда БАВ (например, гистамина, брадикинина, простаглан- дина), которые освобождаются при заболеваниях и травмах легкого. Возбуждение этих окончаний ведет к апноэ, снижению ЧСС и АД, а также к спазму гортани и уменьшению активности скелетной мускулатуры в связи с торможением а-мотонейронов. Это комплексный соматический и висцеральный рефлекторный ответы.
Раздражение рецепторов гортани и трахеи сопровождается кашлем. Чиханье, кашель, смыкание голосовых связок и сужение бронхов, препятствующие попаданию инородных частиц в нижние дыхательные пути, - это защитные рефлексы.
При действии воды на область нижних носовых ходов возникает рефлекс ныряль
щика - рефлекторное апноэ (это тоже защитный рефлекс).
Активация терморецепторов. Сильное возбуждение тепловых или холодовых рецепторов кожи может привести к возбуждению дыхательного центра и усилению дыхания. Однако погружение человека в холодную воду тормозит выдох, и возникает затяжной вдох. Повышение температуры тела при заболеваниях также сопровождается увеличением вентиляции легких. Глубокая гипотермия угнетает дыхательный центр. Незначительное понижение температуры тела стимулирует дыхание.